| 真核生物中的转座元件 | 您所在的位置:网站首页 › satellite of love翻译 › 真核生物中的转座元件 |
真核生物中的转座元件
 转座元件(TE)可以在宿主基因组上自主移动的DNA序列。它们的长度通常在100bp-10000bp之间。和病毒类似,TE也是非常错综复杂并且自私的元件。TE通常能够编码具有多种生化功能的蛋白,或者还含有非编码的调控序列,这些蛋白和调控序列对TE的转座有非常重要的作用。 TE和很多一些类似病毒的侵入性遗传元件的区别并不十分明确。这儿我们定义TE为能够在生殖系细胞中的进行染色体水平重复移动的遗传元件。通过生殖系细胞的移动能够实现TE的垂直传递,以此增加TE的频率。这一定义包含了非自主移动元件,比如短散布核酸元件SINE和微小倒置重复转座元件MITE,同时也包含了内源性逆转录病毒ERV。但是不包括生殖系细胞中来源于病毒的不能整合、移动的内源性元件。此外, 虽然这儿我们定义TE是能够在生殖系细胞中垂直传递的元件,但是要知道,TE的水平转移也是可以发生的。水平转移是TE长期广泛存在的重要原因之一。 对于真核生物而言,除极少数物种外,几乎所有的真核物种基因组中都发现了TE的存在。有些物种中的TE含量可以达到基因组的85%,如果把整个基因组比作一片海洋,那么真正属于宿主的基因可能仅仅是这些海洋中的一些小孤岛。TE在物种中的含量和物种基因组复杂程度无关,有些复杂的多细胞生物(如针叶树、蝾螈等)可以含有较多的TE,但是有些单细胞生物(比如阴道毛滴虫、Anncaliia algerae等)也会含有较多的TE。 在1950年,Barbara McClintock 发表了最早关于TE的论文《The origin and behavior of mutable loci in maize》,她也因此获得了1983年诺贝尔奖。虽然七十余年过去了,人们对TE的研究还存在很多不足。但是人们也认识到TE在物种进化、分化、适应等方面有重要作用。本文综述了在真核生物中,TE的分类、进化起源;不同物种间TE含量的比较;以及影响TE分布的一些因素。 真核生物中TE的分类在1989年,David Finnegan首次将TE划分为两类: Class I:逆转座子Class II: DNA转座子第一类是以RNA为中介,并且将RNA逆转录为DNA,以此整合到宿主基因组中,也就是我们常说的复制粘贴机制。第二类中,大多数是以剪切粘贴机制进行转座,不涉及RNA中介。 根据TE复制、整合等具体过程的不同,上述两类TE可以进一步分成很多子类,或者目order。再往下分,还可以分为超家族、家族等。 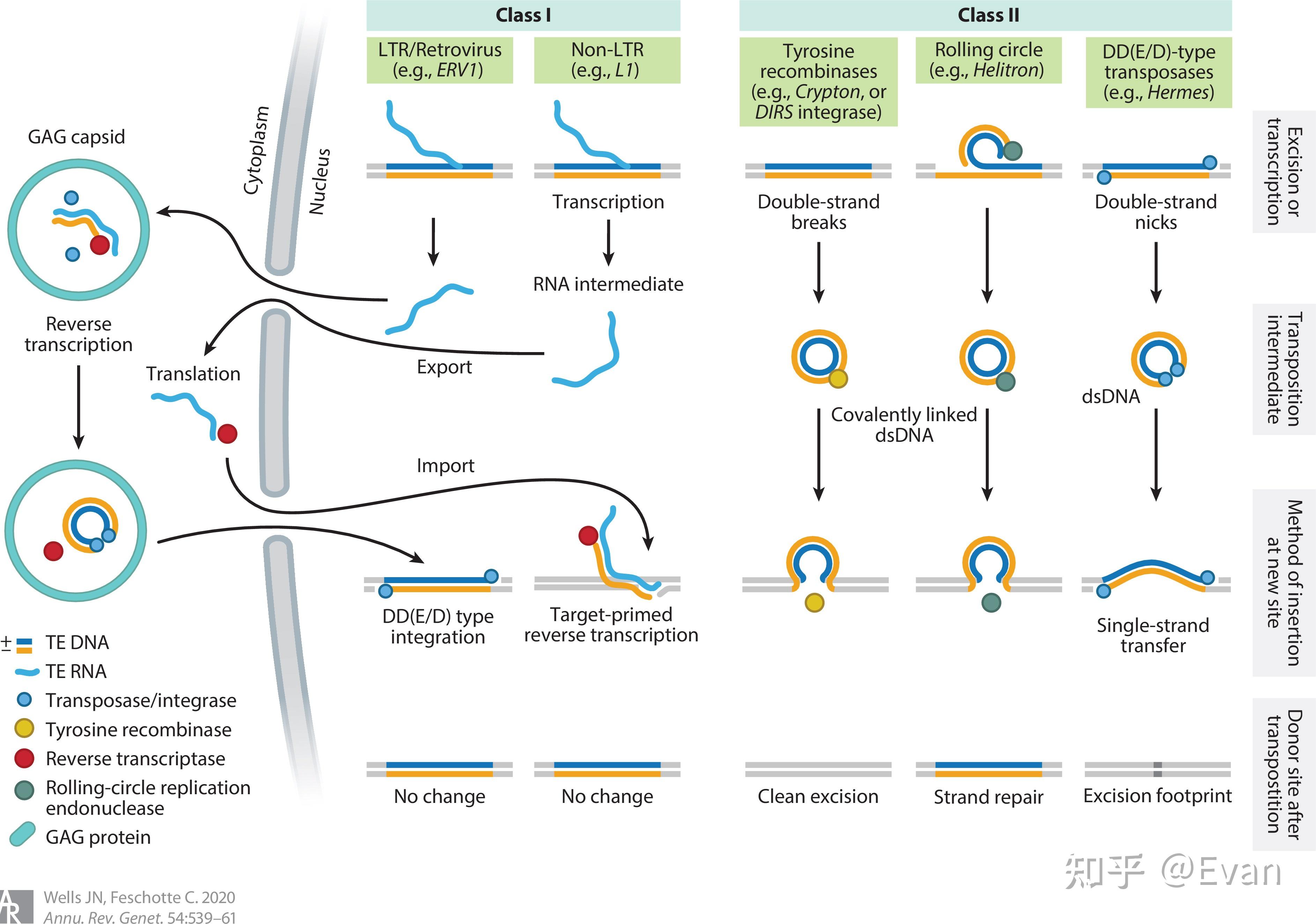 【TE的分类和转座实现方式 - 示意图1】 【TE的分类和转座实现方式 - 示意图1】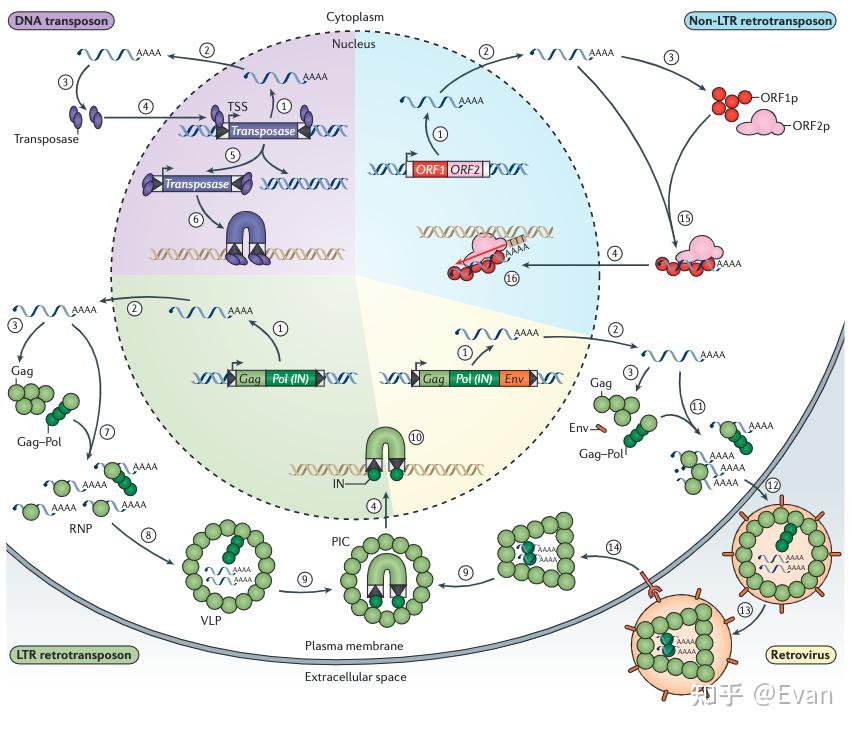 【转座实现方式 - 示意图2】 【转座实现方式 - 示意图2】TE家族的划分一般按照80-80-80方法,即如果两个TE插入的序列长度都超过80bp,有超过80%的序列的相似性超过80%,那么我们可以认为这两个TE是来自同一个家族。因为序列相似性比较高,该家族的TE可以用一个共同的序列来表示该家族共同祖先的序列。特别是当该家族的TE在较短时间内经历了爆发,并且这些TE只经历了中性选择的时候。但是,也有些时候根据80-80-80原则定义的家族和其共同序列并不能够反映各个TE间真实的进化关系。 还有一些TE的分类是根据其能否自主移动划分的。能够自主移动的元件通常都会编码一些移动必需的酶。不能自主移动的元件通常不含相应的编码序列,但是它们能够借助其他可移动元件的顺式序列实现自身的移动。有些TE家族的所有成员都不含有可自主移动的元件,比如MITE。这类TE源于可自主移动的TE,但是其中的自主移动元件在进化过程中被删除了,只剩下了末端倒置重复序列TIR。 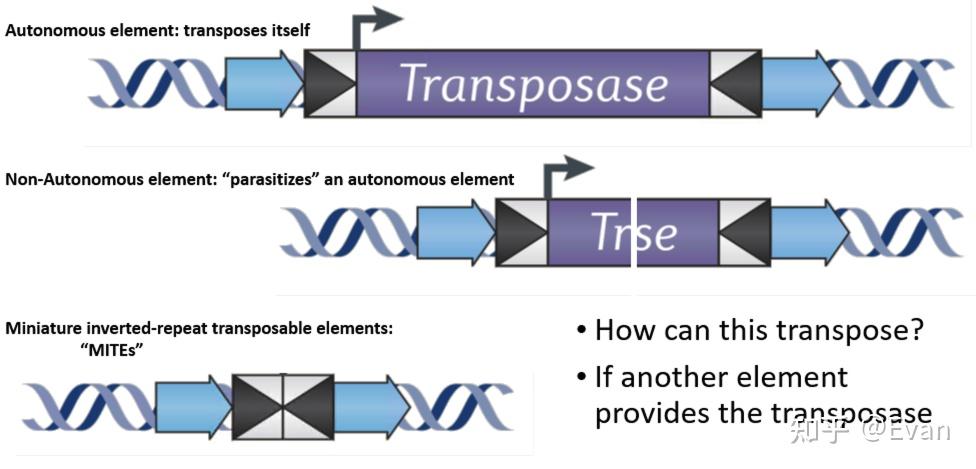 还有一些非自主移动元件(比如SINE)源于非编码基因,比如编码tRNA的基因,其移动需要借助LINE元件的序列。这些必须借助其他家族才能实现自身转座的元件,也被称之为TE的寄生元件,或者是超寄生。 逆转座子根据复制、整合机制的不同,可以将逆转座子分为以下三类 1)长末端重复元件(LTR):比如Copia,Gypsy 2)non-LTR元件:比如L1,R2,Jockey 3)酪氨酸重组酶YR移动元件:比如DIRS Non-LTR的结构比较简单,通常只含有2个开放阅读框(ORF1和ORF2)。其中第一个的开放阅读框ORF1在某些non-LTR群组中缺失(如R2元件)。ORF1的功能可能与RNA的识别和转运有关。第二个开放阅读框ORF2 编码有内切酶EN和逆转录酶RT活性。  【non-LTR元件,R2的第一个开放阅读框缺失,L1等元件则含有第一个开放阅读框。EN:核酸内切酶;RT逆转录酶】 【non-LTR元件,R2的第一个开放阅读框缺失,L1等元件则含有第一个开放阅读框。EN:核酸内切酶;RT逆转录酶】L1元件就是non-LTR家族的代表。在人类基因组中,有数十万拷贝的L1元件,至少有100个拷贝处于活跃状态。其编码的核酸内切酶能够识别剪切5'-TT/AAAA-3'位点,然后合成RNA,翻译逆转录酶,逆转录,最后将逆转录的cDNA整合到宿主基因组中。但是在这个过程中,逆转录常常会提前中断,造成5'端截断。因为5'端是L1表达的重要位点,所以其截断往往会造成L1失活。这也是人类中为什么存在大量的L1却只有相对较少的L1具有活性的原因。 相比于non-LTR,LTR的结构、编码和复制机制比较复杂,并和很多逆转录病毒类似。可自主移动的LTR至少都会含有两个基因:gag和pol;这两个基因编码多聚蛋白,该多聚蛋白能够被pol编码的蛋白酶PR裂解。此外,pol还编码逆转录酶RT,核糖核酸酶HRH,和整合酶IN(下图)。逆转录过程发生在gag形成的病毒样颗粒中,逆转录出的cDNA和整合酶结合,进入细胞核,并整合到宿主基因组中。  【TE分类机及其对应结构】 【TE分类机及其对应结构】逆转录病毒的复制和整合与LTR的转座过程类似。只不过逆转录病毒常常含有env基因。如果逆转录病毒的env基因丢失,那么该逆转录病毒则内源化了。 对YR逆转座元件的研究较少,其结构上与LTR元件相似,但是它编码了酪氨酸重组酶YR,而非整合酶IN。一般认为,YR元件中含有的末端重复序列参与以mRNA为模板的逆转录、单链cDNA的环化、第二条cDNA链的合成,以及最终的整合。 DNA转座子DNA转座子可以分为4类: 1)DDE转座酶介导的剪切粘贴转座:如Tc1/Mariner,P元件 2)酪氨酸转座酶转座子,即Cryptons 3)Helitron 4)Mavericks(也即,Polinton) 其中前两类(DDE和Cryptons)的转座比较简单,结构构成只有一个开放阅读框,编码重组酶,两端含有短末端倒置重复序列(TIRs)。这一结构和很多细菌和古生菌的插入序列很相似。 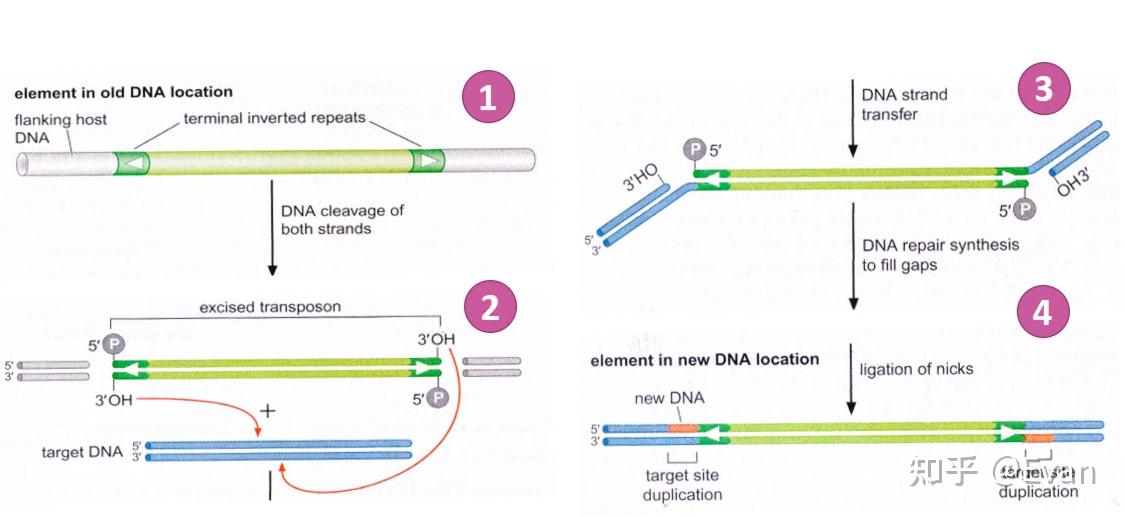 【DDE类转座子结构和转座方式】 【DDE类转座子结构和转座方式】Cryptons在真核生物中分布较少;DDE类转座子是所有TE中分布最广,种类最多的一类转座元件,其至少包含了17个超家族。甚至可以说,DDE是地球上最古老、最丰富的的基因。 不同超家族的DDE转座机制略有差异,但是它们的开始过程都是转座酶对TIR序列的识别和剪切(如上图)。虽然这一个“剪切粘贴”的过程似乎不会增加DDE的拷贝数,但是DDE有一些实现扩增的策略,比如在宿主基因组复制的过程中,DDE可以从已经复制过的位点转座到尚未复制的位点,然后通过宿主基因组的复制,实现自身扩增。此外,在剪切处的双链断裂可以通过同源重组进行修复,修复的过程也能实现一定的扩增。  【DNA转座子通过剪切粘贴实现扩增的方式】 【DNA转座子通过剪切粘贴实现扩增的方式】Helitrons也是一类含量非常丰富的DNA转座元件,在黑腹果蝇、线虫、拟南芥等物种中广泛存在。但是对它的研究仅仅有二十余年的历史。它的结果很简单,没有TIR重复序列等经典DNA转座子结构,不能自主移动,没有“剪切粘贴”。  【Helitrons转座子结构】 【Helitrons转座子结构】Helraiser是其中的典型代表,其通过peel-and-paste机制形成环状双链DNA,然后将此环状DNA转座到基因组其他位置。在转座过程中就实现了扩增。但是在玉米中的研究显示,有些Helitron转座子是通过剪切转座的,而非扩增。所以,对Heliton的转座机制还有待进一步研究。 Mavericks也称之为Polintons,也是一类目前研究尚不完整的DNA转座子。Mavericks转座子的大小约15-20Kb,构成比较复杂,可含有高达20个蛋白编码基因,两端含有400-700bp的TIR重复序列。其在真核生物中也广泛存在,但是其一般在基因组的拷贝数较少(阴道毛滴虫除外,Mavericks可占其基因组的1/3)。 Mavericks和很多双链DNA病毒类似,包括含有pPolB的DNA聚合酶(和腺病毒关系很近),推测其转座过程中可以直接合成DNA,也因此称其为“自我合成型转座子“。Mavericks还含有和DDE类似的整合酶,在整合过程中会形成5-6bp长度的靶标位点重复序列TSD。 有些研究还显示Mavericks可能还能够合成衣壳样蛋白。这提示,Mavericks转座子可能代表了内源性病毒或者噬病毒体。噬病毒体能够嵌合到较大的双链DNA病毒中。Mavericks也因此可能是真核物种为了保护自己免受大DNA病毒攻击的一种秘密武器。 转座元件的进化起源上文描述了各种类型的转座元件,那么这些元件的关系是怎样的,它们是怎么进化的?回答这一问题最好的方法是通过进化基因组学研究各个转座元件的分布和核心蛋白的发生关系。尽管我们现在有很方便的全基因组测序和很好的TE注释工具,但是TE序列进化很快,即便是最常见最保守的TE蛋白域也很难进行超家族之间的比对。此外,TE的进化关系和物种进化关系并不一致,TE可以进行物种之间的水平转移,甚至是在脊椎动物和无脊椎动物之间长距离的转移。有些TE在进化的过程中可能丢失或灭绝了。所以,研究转座元件之间的进化关系非常困难。 在过去数十年的研究中,人们也发现,所有主要转座元件的亚纲subclass在真核生物进化分支中均有分布。对TE核心蛋白的分析也显示这些亚纲在真核生物进化早期就已经存在。同时TE的进化是高度模块化的,可以反复获得或者丢失某些蛋白模块。 转座元件相关蛋白的进化关系尽管各种不同元件的结构非常多样,但是在复制和转座过程中涉及的蛋白种类却很有限,大概可以分为5类: 逆转录酶RT整合酶IN酪氨酸重组酶YRHUH/RepDNA合成酶pPolB而且,HUH、RT和pPolB的还拥有相同的RNA识别域。这些转座涉及的酶可能出现在真核生物形成以前。 在DDE类转座子中,有至少6超家族的转座酶和细菌插入序列IS编码的蛋白相似,这也可能在提示,某些DNA转座子超家族的分化早于真核生物的出现。 除了上述DDE转座子,其他亚纲的转座子和细菌或者古细菌没有同源序列。 在Helitron转座子中,HUH核酸内切酶涉及的转座也出现在细菌IS91的转座过程,但是它们之间应该是独立进化的。HUH核酸内切酶可能源于病毒或者质粒。同样地,在细菌和古细菌中的酪氨酸重组酶和真核生物中转座中涉及的酪氨酸重组酶也应该是独立进化的。这些转座子应该是在真核生物形成之后出现的。 嵌合元件和模块进化上述只描述了相关转座酶之间的进化关系,具体到转座子本身,很难用一个进化树来表示它们之间的进化关系。因为转座子之间会频繁交换结构单元,形成一个进化关系网,而非进化关系树。此外,还有一些辅助结构域是从宿主基因组上获得的。转座子之间的界限并不像物种之间的界限那么明确。 比如,根据逆转录酶结构域,class I中的YR逆转座子可以和LTR及逆转录病毒划分为同一类;但是如果根据酪氨酸重组酶结构,YR逆转座子又可划分为class II中,和Crypton属于一类。 LTR逆转座子是最典型的模块化进化的例子。它从non-LTR和DDE中分别借来了相应的模块,形成了自己独特的转座机制。从进化上看,LTR的形成比non-LTR和DDE要晚,non-LTR和DDE的嵌合形成了LTR。支持这一论断的另一个证据来自核糖核酸酶RNase H结构域,RNase H形成于生命起源的早期,主要作用是降解DNA-RNA复合体中的RNA序列。对LTR、non-LTR和宿主基因组中的RNase H结构域的进化分析显示,LTR的RNase H隶属于non-LTR中的一个分支。如果将宿主基因组的RNase H序列作为树根,那么可以推测出,non-LTR的出现早于LTR。此外,LTR中的gag蛋白和non-LTR中第一个开放阅读框编码的蛋白在序列、位置和功能上都有很多相似之处。最后,LTR中的IN蛋白来自DDE转座子。通过这种模块结构的嵌合,最终形成了LTR转座子。 除了LTR外,SINE类转座子也是通过嵌合模块的方式形式的。比如在灵长类中,Alu在灵长类中通过两个7SL获得的SINE融合得到。Alu形成之后,迅速扩散,并且占领了灵长类基因组转座子的很大一部分。 转座子在物种间分布的差异TE的含量在各个物种间的分布差异很大。有些物种的基因组含有很少的TE,而有些物种基因组中则充斥着大量的TE。  【不同物种的基因组大小和TE含量】 【不同物种的基因组大小和TE含量】有些研究认为,物种中TE的含量和其物种的有效群体数量相关。有效群体数量越大,自然选择效能越高,因而对TE的选择压力也越大。比如,在有效群体数量很大的果蝇中,TE含量较低,而在有效群体数量较小的脊椎动物中,TE插入受到选择压力较小,可以很快的在群体中固定下来。不过,有些有效群体数量相近的物种, 其TE含量有时也会有很大差异。所以有效群体数量的差异并不足解释TE的分布差异。 而且各类不同的TE分布差异也很大,比如LTR在开花植物中分布很多;non-LTR在哺乳动物中分布很多;DNA转座子在斑马鱼和线虫中分布很多。有效群体数量的差异也不足以解释这一现象。 TE丰度和基因组大小除了极少数已知真核生物外(疟原虫、弓形虫、肠脑炎微孢子虫、泰勒原虫),所有的真核生物物种中都含有TE。巧合的是,上述几种例外都是单细胞生物,而且后两者是真核生物中基因组最小的。但是这并不足说明基因组大小和TE丰度有直接关系。另一种单细胞生物,Anncaliia algerae的基因组大小只有23Mb,但是其中有14%都是TE,涵盖了240个TE家族。 在一些较大的基因组中,比如蝾螈基因组,其大小有120Gb。这么大的基因组主要是LTR转座元件造成的。植物基因组通常也可以通过转座元件迅速增大。其中涉及的转座元件可能涵盖较多的TE家族,但是个别TE的作用可能会格外显著。比如棕水螅在3600万年前从绿水螅中分化出来,随后其基因组大小从300Mb迅速增大到了1Gb,造成这一现象的原因就是CR1 non-LTR转座子。 非必要DNA的删除是另一个决定基因组大小和TE丰度的因素。除了转座元件外,蝾螈形成的大基因组和其较低的DNA删除率也有很直接的关系。在拟南芥和水稻中,异位重组造成的基因组高删除率抵消了转座造成的基因组扩大,维持了拟南芥和水稻的基因组大小稳定。在鸟类和哺乳动物中,也有同样的现象。  TE多样性 TE多样性TE在物种之间的分布,除了丰度不同外,种类分布也存在很大差异。宿主和TE之间的竞争作用会导致TE家族结构的形成,扩大TE的亚家族种类(比如L1)。其他一些转座元件,比如Helitrons可以通过获取宿主DNA的片段形成新的亚家族。 不管在什么尺度来衡量,真核生物的TE分布都具有很高的多样性。比如在斑马鱼中,其TE丰度和多样性在脊椎模式生物中都是最高的,含有近2000个TE家族,涵盖了所有的亚纲和几乎所有的超家族。其中,DNA转座子特别丰富,含有1000个不同时期形成的DNA转座子家族,这在鱼类中很不寻常。 但是这并不是说基因组越大,其TE多样性越高。比如云杉是一类裸子植物,其基因组大小20Gb,其中的转座子主要集中在LTR超家族中,含有大量的拷贝数。而且其中的大多数转座子发生在500万-6000万年前。在水稻和玉米中,所有的转座子都晚于500万年。这说明尽管TE在云杉中的多样性很低,但是很多已经存在基因组中的TE会被缓慢的移除掉。与云杉相反,在很多开花植物中,尽管其基因组很小,但是其TE的多样性却很高。甚至在所有陆生植物中,基因组大小和TE多样性还表现出负相关关系。 影响TE分布的因素决定一个TE家族结局的直接因素有三个:1)转座发生率; 2)新TE插入在群体中的固定率;3)TE序列被删除和侵蚀的速率。 影响着三个因素的方面很多,可以概括为:TE自身属性,宿主自身属性。 更广泛地说,TE的命运决定于其自身、宿主和环境三者的相互作用。本部分将重点讨论TE自身因素对其结局命运的影响。 TE插入偏好TE在基因组中插入的位置对TE的结局有重要影响。我们可以通过对自然选择压力进入之前的新插入突变的研究来一窥究竟。研究显示: TE存在一定的插入偏倚TE倾向于在最小化有害作用的基因组中插入TE的靶点倾向于有利于自身增殖TE的插入过程涉及到核酸酶,所有TE编码的核酸酶(限制性内切换、整合酶、转座酶)都在一定程度上存在特定DNA或者染色体属性的特异性,也就是说,几乎所有的TE插入都不是随机的,而是存在一定靶标特异性的。只有一些高度简并性的核酸酶或者识别短序列模式的核酸酶,比如L1,其在人类细胞中的插入才表现出一定的随机性。 很多TE插入表现出很强的位点偏倚,倾向于选择那些不会影响细胞功能的基因组位点进行插入。比如LINE家族的一些TE成员(R1,R2)等,它们的插入精确定位于rRNA基因簇中。因为rRNA基因具有大量的拷贝备份,在其中少量基因的位点的插入不会影响到细胞的整体功能。而对这些位点的识别则是通过高度序列特异性的核酸内切酶实现的。特别是很多类似R2的TE,它们在rRNA基因组的插入也表现出位点特异性,使得它们能够同时存在于基因组中,相互之间不受影响。在多细胞生物中,这种高度特意性的rRNA位点插入是普遍存在的,也证明了TE插入位点选择的偏向性。 基因间的“垃圾”序列为TE的插入和长期存在提供了一个安全的场所。比如在酵母菌中,LTR转座子的插入会避开宿主基因组的基因序列。Copia和Gypsy通过趋同进化,能够选择在Pol-III转录基因序列的上游插入,避免干扰宿主基因表达。Ty5能够通过其整合酶和一些染色质因子的作用实现其在端粒区非表达染色质的特异插入。 很多TE的插入靶标位于基因5'端的上游,这种靶标选择倾向会给TE自身带来益处。首先在该区域的插入能够避免插入对编码蛋白的干扰,同时这些区域的染色体多以染色质形态存在,有利于TE自身的表达和转座。很多DNA转座子都采用了这种策略来实现自身利益最大化,比如果蝇中的P元件、玉米中的MuDR,大米中的mPing和拟南芥中的VANDAL21等。 酵母菌中的Tf1逆转座子同样倾向于选择基因的启动子区域插入,该转座子的整合酶和宿主特异性转录因子相互作用,实现插入位点的选择。 在拟南芥和其他一些植物中,类Copia的逆转座子也进化出了一些机制,实现在宿主非必需基因中插入。这些机制主要是通过识别核小体组蛋白H2A.Z来实现的,该组蛋白不存在于必需基因组中,只存在于和适应环境压力相关的非必需基因中。这也提示,TE非随机插入带来的基因组突变可能也有利于宿主适应外界环境的变化。 还有一种TE的插入策略比较特殊,它们倾向于选择其他的TE序列作为自己的插入靶点。研究显示有些TE家族会频繁地出现在其他TE家族的序列内。比如在非洲爪蟾中,non-LTR转座子Tx1L几乎只存在于DNA转座子Tx1D中,前者像是后者的一个寄生虫,这种现象可以称之为TE的“超寄生”hyperparasitism. 影响TE长期存在的因素所有的新TE插入都会受到来自宿主水平的自然选择。特别是当TE对宿主产生有害作用时,比如:干扰到宿主基因的表达;TE表达产物对宿主细胞有毒副作用;同家族TE导致的宿主染色体异位重组。  【TE导致的人类疾病】 【TE导致的人类疾病】虽然有些TE产物确实对宿主有害,比如L1转座中间产物的积累会导致Aicardi-Goutières综合征,但是很多时候毒性并不是主要受选择的因素。目前的研究显示,TE导致的异位重组是限制TE增殖的主要因素。 序列较长的TE更容易造成异位重组,因而其受到的选择压力也应该更大。实验观测也确实如此,比如LTR和LINE等较长的转座子常常会聚集在低重组区(中心粒周围的异染色质区),在这些区域内,TE受到的选择压力会相对较小。与之相反,一些较短的转座子,比如SINE和MITE,通常富集在基因较多的染色体区域,这些区域通常重组率也比较高。 第二个导致TE受到选择的因素是其对基因表达的影响。可自主移动的转座子其自身通常会含有启动子和调节序列,如果其插入到基因序列中,那么对宿主基因的表达会产生较大影响。比如,在人类基因组中,L1转座子很少出现在基因序列中,比较老的LTR插入也很少出现在基因序列两侧5kb的的范围内。这都证明了携带启动子的LTR在人类基因组中受到很强的自然选择压力。即便有些插入出现在基因中,也常常是内含子中,而且是内含子的中段部分,以尽量避免对外显子的影响。 当然,还要说明一点,TE并不一定只给宿主带来坏处,还有可能给宿主带来适应优势,比如果蝇中Doc(non-LTR)的插入导致了Cyp6g1基因表达增加,该基因提高了宿主对DDT等杀虫剂的抗药性。 TE水平转移有性生殖为TE的垂直传递提供了便利,但是TE的水平转移也是其长期存在的重要原因。几乎所有的TE都存在水平转移现象,在某些TE家族会表现的特别明显。对于DDE类型的DNA转座子,其水平转移相对容易,发生率也更高;而相对于non-LTR的逆转座子,其水平转移比较罕见。 DNA转座子(DDE)容易发生水平转移可能是因为其进化出了一些转座机制,最大程度上减少了宿主特异性。比如,它们会编码一些弱作用的启动子,甚至不编码启动子,这使得它们的转座必须依赖于插入到宿主基因表达调节序列中,利用宿主的启动子实现转座,其中涉及到的很多转录因子也是直接使用宿主提供的。这大大提高了他们在不同宿主中的适应性。此外,有些DNA转座子的转座酶在不同宿主细胞中都能保持较高活性,甚至在非细胞的体外环境也能保持一定的活性。这些因素都降低了DNA转座子宿主的特异性,促使其水平转移更加容易。 转座中介物的不同也是造成TE发生水平转移差异的原因。比如转座中的DNA中介比RNA中介产物更加稳定,DNA转座子更容易水平转移。有些逆转座子能够形成类似病毒可以的衣壳,这些衣壳会促进这些逆转座子在细胞外的播散。有些转座子可以依赖真正的病毒进行水平转移,比如转座过程中的中介物可以与病毒序列结合或者利用病毒衣壳。中介物产生的位置会影响水平转移,比如有些中介物主要(non-LTR)或者局限(DNA转座子)在细胞核中,那么这些转座子可能很难依赖在细胞质中进行自我复制的病毒(如痘病毒)而进行水平转移;不过它们却可以依赖一些在细胞核中复制的病毒(比如疱疹病毒和杆状病毒)进行水平转移。 抵御宿主免疫宿主也会对TE活动进行抑制,特别是在生殖系细胞中。Piwi(piRNA)在生殖细胞中很活跃,通过RNA干扰,能够抑制TE的活性。比如在黑腹果蝇的卵子形成过程中,I元件在卵母细胞中进行逆转座,但是其RNA中介物仅仅在卵母细胞周围的支持细胞中生成,然后等卵母细胞成熟之后,再通过微管系统从支持细胞转运至卵母细胞中。这么做的目的一方面是周围支持细胞数量较多,能够积累很高浓度的RNA中介物;另一方面也是避免卵母细胞中piRNA的抑制作用。 总结转座子(TE)几乎存在于所有的物种中,对物种的基因组结构和物种进化有重要的影响。尽管如此,人们对TE的研究还相当欠缺。在发现TE的前60年的时间内,人们主要是通过模式生物和少数农业作物对少数TE进行描述,研究其对表型的影响。最近十余年,基因组测试技术的进步使得我们能够更加全面的识别、比较、探究不同TE的调节、转录活性,对TE的研究更加具体详细。人们发现基因组中大部分TE都是失活的转座遗迹,犹如一座座死火山,这些遗迹的累积构成了真核生物基因中的重复序列。 对TE的研究不能忽视对其转座机制的研究。目前越来越多的研究出现在了基因组进化水平上的研究,却忽略了TE本身的转座机制。 TE家族之间表现出了很大的差异性,其对宿主基因组造成的影响也很大。在更多的物种中,对TE进行识别和分类也同样重要。同时要注重对转座机制的研究,特别是一些比较特殊的转座子,比如Helitrons, Marvericks, YR元件等。 参考文献: Wells, J. N., & Feschotte, C. (2020). A field guide to eukaryotic transposable elements. Annual review of genetics, 54, 539-561. Chenais et al (2013) Biochimica et Biophysica Acta 1835 (2013) 28–35,Transposable elements and human cancer: A causal relationship? Sultana, T., Zamborlini, A., Cristofari, G., & Lesage, P. (2017). Integration site selection by retroviruses and transposable elements in eukaryotes. Nature Reviews Genetics, 18(5), 292-308. |
【本文地址】